Nutrigenomics helps to understand better the cellular mechanisms influencing on physiological and metabolic processes relevant for animal production, such as lipid metabolism (Benítez et al., 2017). It involves multi-disciplinary fields, such as nutrition, bioinformatics, molecular biology, genomics, functional genomics, epidemiology, and epigenomics to study the complex interactions of the genome and the diet (Benítez et al., 2017).
The amount of intramuscular fat or marbling, for example, is an important quality characteristic due to its effect on meat flavor and juiciness; and some fatty acids in the intramuscular fat, such as conjugated linoleic acid (CLA, c9, t11-C18:2) and long chain n-3 fatty acids can be relevant to increase for their role in human nutrition (Bessa et al., 2015).
The interactions between nutrients and genes start with the maternal influences on the offspring gene expression (fetal programming), where nutrient restriction or excess diet in the late gestation and neonate phases can have long-term effects (Ladeira et al., 2018). Another key moment of these interactions is around 150 days until 250 days of life, where adipogenesis in the intramuscular fat is still occurring but not in other fat depots (marbling window). In that period it can be applied a strategy to increase in the number of adipocytes in intramuscular but not subcutaneous, internal or visceral fats.
However, marbling will be only visible if the existing adipocytes have grown during the hypertrophy period, so the dietary energy in later stages needs to be high. In the fattening phase triglycerides are incorporated into the adipocytes, after the absorption of dietary fatty acids or by de novo synthesis of fatty acids (Ladeira et al., 2016).
Acetate and glucose are the major precursors used for biosynthesis of fatty acids in ruminants, where intramuscular adipocytes prefer glucose, and subcutaneous adipocytes prefer acetate as lipogenic substrates (Ladeira et al., 2018). Therefore, enhancing glucose by feeding cereals/concentrates and reducing roughages can promote marbling but not back fat deposition. Furthermore, delta-9 desaturase activity (SCD) is considered a key regulatory enzyme of adipogenesis and high grain diets increase SCD activity and intramuscular fat deposition (if low rumen pH is avoided, Teixeira et al., 2017).
However, the development of nutritional strategies to increase SCD activity and adipogenesis in forage fed ruminants is a major goal for research. If this can be achieved by somehow simulating the hyper-insulinemic, or by increasing muscle insulin sensitivity, or by other approaches remains unsolved (Bessa et al., 2015).
Dietary fats provide energy, affect the ruminal microbiota and deliver particular fatty acids whose effect on lipid metabolism can be differentiated, affecting both the amount of intramuscular fat and their fatty acid composition. Mono-unsaturated fatty acids and polyunsaturated fatty acids in general cause down-regulation of SCD1 expression to discourage de novo lipogenesis (Bessa et al., 2015; Ladeira et al., 2018).
Typically beef from forage diets could be claimed as source of long chain n-3 fatty acids (Pethick et al., 2021). From recently published research, fifteen showed an improvement in the fatty profile by animal diets, including the concentrations of CLA and oleic acid; and decrease hypercholesterolemic fatty acids and n-6/n-3 ratio (Ladeira et al., 2018). Regarding concentrate diets, the rumen outflow of t11-18:1 frequently is replaced by t10-18:1 (t10-shift) which cannot be converted to CLA, which makes it difficult the increase of this fatty acid in the meat, relevant for human health (Bessa et al., 2015).
The knowledge on the cellular mechanisms influencing the amount of intramuscular fat and their composition may allow nutritionists to use feedstuffs and additives that modulate the expression of target genes, aiming at the improvement in meat quality (Ladeira et al., 2018). The complexity of ruminant metabolism, determined mainly by its symbiotic rumen microbiota, will continue to challenge the efforts to modulate meat quality including its fatty acid composition (Bessa et al., 2015).
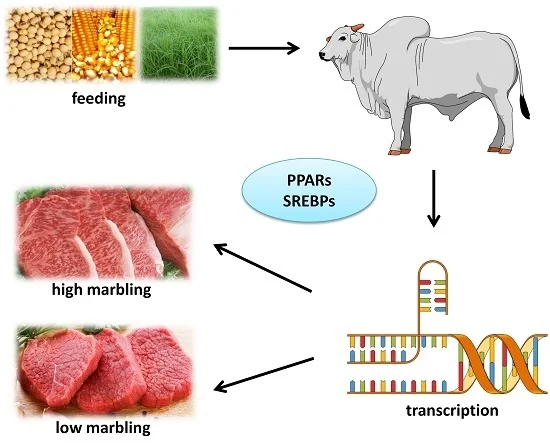 Image 1. Graphical abstract from Ladeira et al. (2016).
Image 1. Graphical abstract from Ladeira et al. (2016).
Authors: Virginia C. Resconi and Jakeline Vieira Romero (Universidad de Zaragoza, Spain).
 | |
 | Appropriate nutritional management prevents clinical and subclinical diseases. |
 | A challenge is to find nutritional practices to achieve simultaneously a meat more nutritive and palatable, without negative effects on the productive performance. |
 |
Source of information
- Ladeira, M.M. et al. (2018). Animal. 12:s282-s294. https://doi.org/10.1017/S1751731118001933
- Ladeira, M.M. et al. (2016). International Journal of Molecular Sciences. 17: 918. https://doi.org/10.3390/ijms17060918
- Benítez, R. et al. (2017). Journal of Investigative Genomics. 4:23-28. https://doi.org/10.15406/jig.2017.04.00059
- Rui, J.B. et al. (2015). European Journal of Lipid Science and Technology. 117:1325-1344. https://doi.org/10.1002/ejlt.201400468
- Pethick, D.W. et al. (2021). Animal. 15: 100356. https://doi.org/10.1016/j.animal.2021.100356
Further information
- Bordoni, L. & Gabbianelli, R. (2019). Biochimie. 160:156–171. https://doi.org/10.1016/j.biochi.2019.03.006
- Nowacka-Woszuk, J. Animal Genetics. 61:93-103. https://doi.org/10.1007/s13353-019-00522-x
- Önenç, S.S. & Ozdogan, M. (2022). Journal of Animal Production. 63:67-74.
https://dergipark.org.tr/en/download/article-file/1163488
- Orrù, L. et al. (2011). Meat Science. 87:344–348. https://doi.org/10.1016/j.meatsci.2010.11.009
- Waters, S.M. et al. (2009). Journal of Animal Science. 87:244-252. https://doi.org/10.2527/jas.2008-1005